CAILE
PE CARE SE REALIZEAZA VARIATIA GENETICA SI VARIABILITATEA
FENOTIPICA IN POPULATIILE DE MICROORGANISME, PLANTE SI ANIMALE
Variabilitatea
fenotipica a indivizilor ce formeaza populatiile de
microorganisme, plante si animale, este reflectarea polimorfismului
genetic existent la nivelul lor, adica a variatiei genetice
inter-individuale sau intrapopulationale. Aceasta variatie este
rezultatul recombinarii genetice, mutatiilor si activitatii elementelor genetice
transpozabile.
Recombinarea
genetica
Organismele vii se
caracterizeaza, printre altele, prin faptul ca intre indivizii unei
populatii, respectiv ai unei specii, exista nenumarate
deosebiri, atat genotipice, cat si fenotipice.
Fiecare
individ constituie o entitate unica si nerepetabila (exceptand
indivizii clonati), datorita faptului ca el prezinta o
anumita configuratie a factorilor genetici.
Deosebirile
genotipice dintre indivizi se realizeaza in primul rand prin fenomenul
recombinarii genetice, prin care informatia ereditara
are capacitatea de a "circula" in cadrul unei populatii.
Recombinarea se poate realiza pe parcursul meiozei, caz in care este
denumita recombinare meiotica,
dar poate sa se produca si in celulele care parcurg mitoza, caz
in care poarta denumirea de recombinare
mitotica sau somatica.
La
speciile evoluate, frecventa recombinarii mitotice este foarte
scazuta comparativ cu aceea a recombinarii meiotice, dar in
ambele tipuri de recombinare procesul este reciproc, constand in schimbul
anumitor gene intre un grup linkage si grupul omolog din cromozomul
pereche.
Fenomenul
recombinarii genetice are o importanta biologica
extraordinara, acesta determinand diferentierile genotipice intre
indivizii unei populatii sau ai unei specii. Variatia genotipica
reprezinta suportul pentru caracterul unic al indivizilor, pentru
asigurarea unor posibilitati largi de adaptare la variatele
conditii de mediu si, implicit, pentru asigurarea supravietuirii
populatiilor sau speciilor.
Recombinarea
genetica se realizeaza in principal in cadrul procesului sexual, fapt
care explica raspandirea sa extraordinara nu numai la
organismele evoluate, dar chiar si la procariote, cum sunt de exemplu
bacteriile.
La
organismele eucariote, fenomenul de recombinare genetica se
realizeaza, in cadrul procesului sexual, pe trei cai:
prin disjunctia
independenta a perechilor de cromozomi in cursul meiozei (recombinare inter-cromozomiala);
prin crossing-over intre cromozomii pereche (recombinare
intra-cromozomiala);
prin conversie (recombinare genetica nereciproca).
Disjunctia independenta a perechilor de cromozomi
(recombinarea inter-cromozomiala)
Studiul comparativ al
mitozei si meiozei a aratat ca al doilea tip de diviziune este
cel care asigura libera combinare a cromozomilor si, pe aceasta
baza, o mare variatie genotipica a gametilor si a
descendentilor.
Cromozomii
omologi care se asociaza in metafaza I formand bivalenti, se
separa ulterior, fiecare pereche independent de celelalte, fapt care
determina combinarea pe baza de probabilitate a cromozomilor
proveniti de la bunici. In mod logic, posibilitatile de a forma
mai multe combinatii in cadrul gametilor sunt direct corelate cu
numarul de perechi de cromozomi.
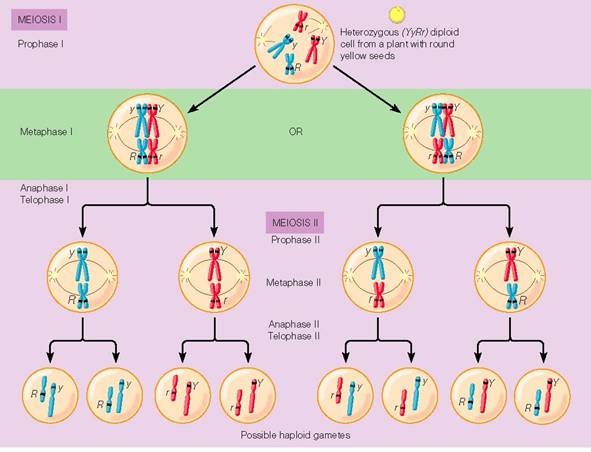
RECOMBINAREA
INTERCROMOZOMIALA IN CURSUL MEIOZEI
Spre exemplu, daca se incruciseaza doua specii care au
2n=6, si daca pe
fiecare pereche de cromozomi se afla cate o singura pereche de gene,
hibridul care rezulta va mosteni jumatate din cromozomii si
factorii ereditari ai fiecaruia dintre genitori. Astfel, daca
genitorul matern poseda genele AA BB
CC si cel patern genele aa bb cc,
hibridul va fi Aa Bb Cc.
In
cursul meiozei, acest hibrid va forma 8 tipuri de gameti datorita
faptului ca in metafaza primei diviziuni meiotice (diviziunea
reductionala) cromozomii omologi se grupeaza in bivalenti
(cate doi) si apoi migreaza la polii celulei in mod diferit,
indiferent de originea lor.
Rezultatul
este formarea a 8 tipuri de gameti masculi si femeli diferiti
genotipic (ABC, ABc, AbC, Abc, aBC,
aBc, abC, abc), prin a
caror unire la intamplare vor rezulta 64 de combinatii genetice
diferite si respectiv 8 tipuri de descendenti, diferiti din
punct de vedere al fenotipului.
|
♀ ♂
|
ABC
|
ABc
|
AbC
|
aBC
|
Abc
|
aBc
|
abC
|
abc
|
|
ABC
|
AABBCC
|
AABBCc
|
AABbCC
|
AaBBCC
|
AABbCc
|
AaBBCc
|
AaBbCC
|
AaBbCc
|
|
ABc
|
AABBCc
|
AABBcc
|
AABbCc
|
AaBBCc
|
AABbCc
|
AaBBcc
|
AaBbCc
|
AaBbcc
|
|
AbC
|
AABbCC
|
AABbCc
|
AAbbCC
|
AaBbCC
|
AAbbCc
|
AaBbCc
|
AabbCC
|
AabbCc
|
|
aBC
|
AaBBCC
|
AaBBCc
|
AaBbCC
|
aaBBCC
|
AaBbCc
|
aaBBCc
|
aaBbCC
|
aaBbCc
|
|
Abc
|
AABbCc
|
AABbcc
|
AAbbCc
|
AaBbCc
|
AAbbcc
|
AaBbcc
|
AabbCc
|
Aabbcc
|
|
aBc
|
AaBBCc
|
AaBBcc
|
AaBbCc
|
aaBBCc
|
AaBbcc
|
aaBBcc
|
aaBbCc
|
aaBbcc
|
|
abC
|
AaBbCC
|
AaBbCc
|
AabbCC
|
aaBbCC
|
AabbCc
|
aaBbCc
|
aabbCC
|
aabbCc
|
|
abc
|
AaBbCc
|
AaBbcc
|
AabbCc
|
aaBbCc
|
Aabbcc
|
aaBbcc
|
aabbCc
|
aabbcc
|
Cu cat este mai mare numarul cromozomilor si respectiv al genelor
in cromozomi, cu atat este mai mare numarul combinatiilor posibile
ale gametilor. Astfel, daca in cazul unei specii cu 8 cromozomi
si cate o singura gena in cromozom, probabilitatea ca un gamet
sa fie diferit de altul este egala cu (1/2)4 = 1/16, iar
ca un individ sa fie diferit de altul este in consecinta de
(1/2)8 = 1/256, la o specie cu 20 de cromozomi probabilitatea ca un gamet sa fie
diferit de altul este de (1/2)10 = 1/1.024, iar ca un individ
sa fie diferit genotipic este de (1/2)20 = 1/1.048.576.
Daca am considera ca la specia luata in considerare sunt numai
trei gene, plasate una de alta la o distanta suficient de mare,
astfel incat sa se poata separa usor prin crossing-over, probabilitatea
ca un individ sa fie diferit genotipic de altii ar fi de (1/2)60.
Calculul matematic arata ca, pentru a oferi spatiu tuturor
combinatiilor genotipice posibile intr-un astfel de caz, ar fi
necesara o suprafata de teren de 2.000 ori mai mare decat cea a
Pamantului.
Inmultirea sexuata a organismelor
determina asadar o mare variabilitate genotipica a
descendentilor, in primul rand prin mecanismul de combinare pe baza
de probabilitate a cromozomilor in cursul diviziunii reductionale (prima
diviziune meiotica).
Pe
langa aceasta, un rol important in sporirea variatiei genotipice
individuale revine fenomenului de schimb de segmente cromatidice intre
cromozomii pereche (crossing-over) precum si genelor extranucleare.
Recombinarea genelor intre cromozomii
pereche (crossing-over)
Studiul mecanismului de transmitere
ereditara a aratat ca nu intotdeauna genele care fac parte din
aceeasi grupa de linkage, deci plasate in acelasi cromozom, se
transmit inlantuite, si ca de la acest mod de transmitere a
informatiei genetice exista unele exceptii. Respectivele abateri
au fost explicate prin posibilitatea realizarii unui schimb reciproc de
gene intre cromozomii pereche, printr-un proces care a fost denumit crossing-over.
Efectul
recombinarii intra-cromozomiale este disocierea dintr-o legatura
a unor gene si formarea unei noi grupe linkage cu genele cromozomului
omolog.
Frecventa cu care
se poate realiza recombinarea prin fenomenul de crossing-over este foarte
redusa si numai in cazuri rare se apropie de limita superioara
de 50%, valoare care corespunde segregarii independente a caracterelor,
conform legilor mendeliene.
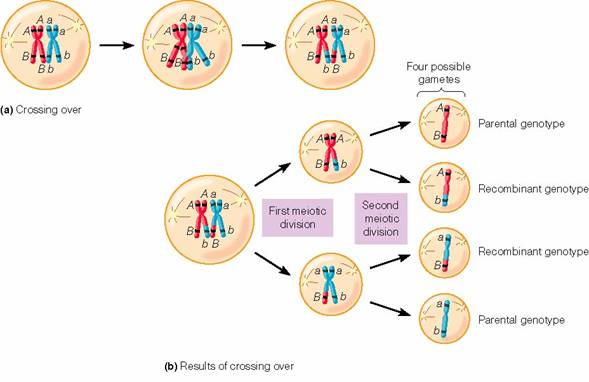
Crossing - over (schimbul de
segmente de cromatide intre cromozomii pereche)
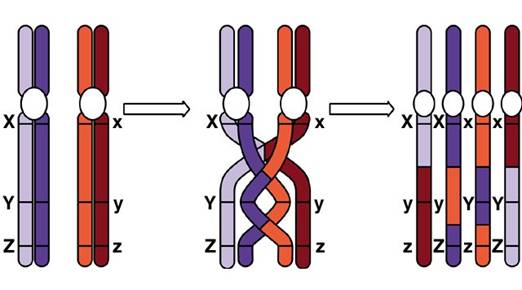
Crossing - over in cursul meiozei
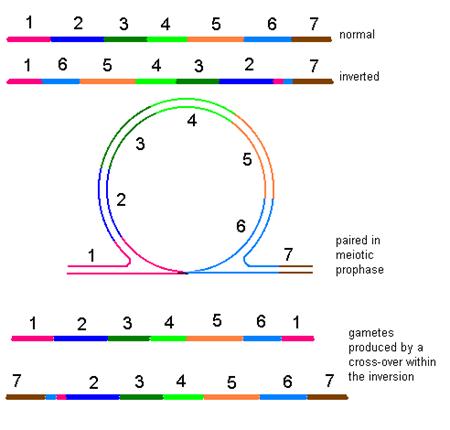
Un tip
particular de crossing over, realizat in cadrul unei inversii
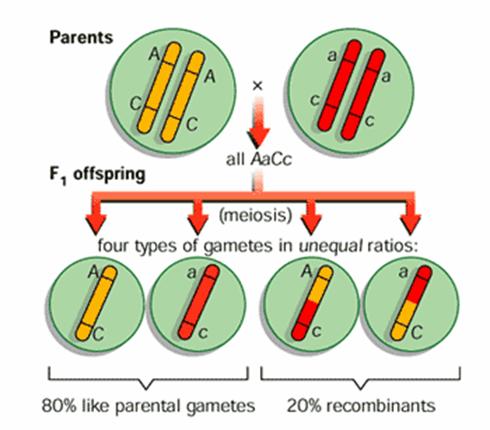
Formarea
de gameti recombinati - baza a polimorfismului
genetic
Recombinarea
genetica nereciproca (conversia genica)
Spre deosebire de recombinarea
genetica reciproca, care se realizeaza prin crossing-over,
recombinarea genetica nereciproca apare ca rezultat al fenomenului de
conversie.
Conversia se manifesta prin aceea
ca descendenta haploida a nucleului heterozigot diploid (a+
a) nu prezinta raportul normal de segregare, respectiv 1:1, una dintre
alele avand o frecventa mai mare decat cealalta. Astfel, la
ciuperci, in locul raportului de segregare normal 4:4 se intalnesc raporturile
de 5:3; 6:2; 7:1; 8:0. S-a demonstrat ca frecventa recombinarii
genetice nereciproce variaza la diferite gene, iar in interiorul unei gene
frecventa conversiei creste de la un capat la altul. Fenomenul
de polarizare a conversiei in interiorul aceleeasi gene poarta
denumirea de polaron.
Fenomenul
de conversie are loc frecvent la bacteriile lizogene, care contin profagi
si din aceasta cauza dobandesc caracteristici noi privind
producerea de toxine, sensibilitatea la infectii cu alte virusuri, sau
isi modifica particularitatile antigenice. Acest tip de
conversie se numeste conversie
fagica si este cel mai probabil cauzata de aditia de
material genetic la genomul bacterian.
Conversia genica are loc in cursul
meiozei si se considera ca se realizeaza in doua
etape:
In prima etapa are loc transferul a 100-200 nucleotide din molecula de
ADN de pe o cromatida, pe alta, formandu-se astfel scurte secvente de
ADN hibrid;
In cea de a doua etapa are loc corectia imperecherilor
gresite dintre nucleotide in cadrul ADN hibrid, in timpul sau imediat
dupa recombinarea intergenica prin crossing-over. Ca urmare s-a observat existenta unei corelatii intre
fenomenul crossing-over si frecventa conversiei in imediata
vecinatate.
MUTATIILE
Mutatia
defineste orice modificare
ereditara si detectabila a materialului genetic, care nu este
cauzata de recombinarea genetica sau de segregare. Mutatiile
cauzate de factori de mediu (fizici sau chimici) sau de factori biologici apar
spontan si sunt denumite mutatii naturale. In
functie de materialul genetic afectat, mutatiile sunt clasificate
astfel:
a)
mutatii genice, cand sunt afectate genele;
b)
mutatii cromozomiale, cand sunt afectati
cromozomii;
c)
mutatii genomice, cand este afectat intregul
genom (mutatii
multiple, la nivelul mai multor
perechi de cromozomi).
In
functie de genele care sunt afectate, implicand deci exprimarea
fenotipica, mutatiile pot fi clasificate in mutatii dominante, co-dominante, semidominante
si recesive.
Mutatiile care se produc la
nivelul perechilor de nucleotide sunt denumite mutatii punctiforme si sunt evident mutatii
intragenice.
Mutatiile au
fost de asemenea clasificate si in functie de locul unde sunt plasate
genele afectate, respectiv pe autozomi sau heterozomi, in mutatii autozomale si mutatii heterozomale (care manifesta sex-linkage).
Exista si mutatii ale genelor din
citoplasma, denumite mutatii extranucleare.
O categorie relativ distincta
de mutatii o constituie mutatiile
letale si mutatiile semiletale,
care afecteaza gene de importanta majora in organism, prin
a caror blocare se cauzeaza moartea individului, cel mai adesea in
perioada embrionara, sau inainte de maturitatea sexuala.
Indiferent de
nivelul la care se produc (genic, cromozomial, genomic), mutatiile au
frecventa variabila. Se apreciaza
insa ca pot exista gene mutabile, caracterizate printr-o mare
instabilitate si care, in consecinta, prezinta o
frecventa mai mare a mutatiilor, comparativ cu celelalte gene
din organism. O astfel de ipoteza este de
altfel sustinuta de existenta seriilor de gene polialele, care
sunt considerate ca fiind rezultatul instabilitatii pronuntate a
unor gene sub influenta anumitor conditii ale mediului abiotic. S-a sugerat de asemenea posibilitatea existentei unor gene mutatoare, care maresc
frecventa mutatiilor altor gene.
Mutatiile naturale si frecventa lor
Se considera ca una dintre
cauzele mutatiilor naturale care apar la organismele vegetale si
animale este radiatia cosmica, o sursa permanenta de nuclei
de carbon, azot si oxigen, care ciocnindu-se cu nuclei ai unor atomi din
aer dau nastere radiatiei cosmice secundare, formata din
radiatii electromagnetice si corpusculare. Aceste radiatii
actioneaza permanent asupra Pamantului, impreuna cu
radiatiile care se produc prin dezintegrarea naturala a elementelor radioactive din scoarta pamantului
si cu gazul radon din atmosfera, alcatuind fondul natural de
radiatii terestre.
Atunci cand asupra organismelor
dintr-o populatie actioneaza un factor
de stres fizic sau chimic, respectiv un factor mutagen, probabilitatea
producerii unor mutatii este semnificativ mai mare si, de
regula, are loc o crestere a frecventei mutatiilor.
Mutatiile naturale au o
frecventa foarte variata in functie de conditiile de
mediu si de genele afectate. Frecventa mutatiilor se poate
calcula numai pentru o anumita gena, dar insumand valoarea frecventei
mutatiilor pentru toate genele se poate estima frecventa
mutatiilor per genom.
Rata
mutatiilor nu este constanta, ci depinde de diferiti factori,
printre cei mai importanti fiind genotipul si conditiile de
mediu. Genotipul poate influenta intr-o masura foarte
importanta rata mutatiilor, mai ales la acele organisme la care s-a
demonstrat existenta asa-numitelor gene mutatoare, care maresc frecventa mutatiilor la
diferite alte gene. Cauzele posibile ale activarii la nivel molecular a mecanismului de
actiune al genelor mutatoare pot fi:
a) prezenta unei polimeraze anormale, care determina erori in replicatia ADN;
b) producerea unor analogi ai bazelor azotate cu caracter mutagen, care in
cursul replicatiei ADN sunt inclusi in macromolecula respectiva;
c) modificarea unor baze azotate din macromolecula de ADN, avand ca rezultat
aparitia de erori in cursul replicatiei.
Descoperirea
unei gene care codifica producerea unei polimeraze anormale la organisme
caracterizate printr-o frecventa marita a mutatiilor
reprezinta o dovada in sprijinul acestei ipoteze.
Factorii Mutageni
Faptul ca radiatiile pot
induce mutatii este cunoscut inca din anul 1927, cand H.J. Muller a
demonstrat cresterea semnificativa a frecventei mutatiilor
induse artificial in raport cu aceea a mutatiilor spontane. Ulterior s-a
descoperit ca numeroase substante chimice (compusi chimici
naturali sau de sinteza) au efect mutagen. S-a demonstrat apoi ca
socurile termice pot de asemenea induce aparitia de mutatii la
unele organisme, prin favorizarea instabilitatii materialului genetic
si a replicarii sale eronate.
Factorii
mutageni pot fi clasificati asadar in:
factori mutageni fizici;
(2) factori mutageni chimici;
(3) factori mutageni biologici.
Indiferent de categoria
din care fac parte, actiunea constanta sau prelungita a
agentilor mutageni determina a marire considerabila a
frecventei mutatiilor.
Factorii mutageni fizici
In grupa factorilor mutageni fizici
sunt incluse radiatiile ionizante si neionizante, precum si
socurile de temperatura. Radiatiile care pot provoca
mutatii la nivelul diferitelor unitati ale materialului genetic
(gene, cromozomi, genom) sunt clasificate astfel:
Radiatii neionizante (care
genereaza reactii fotochimice):
-
raze ultraviolete (UV)
Radiatii
ionizante (care genereaza reactii radiochimice):
Radiatii electromagnetice:
-
raze X (Röntgen)
-
raze gamma (g
Radiatii corpusculare
-
raze beta (electroni)
-
protoni (nuclei de hidrogen)
- neutroni lenti sau rapizi
-
raze alfa (nuclei de heliu)
- particule grele
Radiatiile neionizante sunt
radiatiile ultraviolete (UV), care fac parte din spectrul solar invizibil,
fiind constituite din fotoni cu energie joasa (circa 3-5 ergi/m2)
si cu o lungime de unda cuprinsa intre 136 si 4.000 Å.
Actiunea radiatiilor UV se
exercita prin absorbtia energiei fotonilor cu diferite lungimi de
unda de catre moleculele substratului, care intra intr-o stare
de excitatie. Consecinta acestei stari este aparitia unor
reactii chimice fluorescente si fosforescente, si transmiterea
energiei moleculei excitate unei alte molecule. Asemenea fenomene provoaca
reactii chimice secundare variate in functie de substratul asupra
caruia se exercita actiunea si de lungimea de unda a
razelor UV.
Radiatiile ionizante includ radiatiile electromagnetice si
radiatiile corpusculare, care au
un efect similar asupra substratului. Acestea se exercita ca unde sau ca
particule incarcate cu energie diferita si actioneaza
la nivelul atomilor, fiind capabile sa dezorganizeze sferele de electroni
care inconjoara nucleul atomic.
Se
stie ca atomul neutru este intr-un echilibru electrostatic,
numarul sarcinilor electrice pozitive din nucleul atomic fiind egal cu cel
al sarcinilor electrice negative ale electronilor.
Radiatiile electromagnetice,
datorita energiei si a vitezei lor foarte mare, sunt capabile sa
smulga unul sau mai multi electroni de pe orbita exterioara a
atomilor neutri intalniti in drumul lor. Prin pierderea unui electron,
atomul ramane cu o sarcina electrica pozitiva in plus,
devenind ion pozitiv sau cation. Electronul eliberat nu ramane liber, ci se
ataseaza unui alt atom care, primind o sarcina electrica
negativa, devine ion negativ sau anion. Acest proces poarta numele de
ionizare. Desi ionii
produsi in urma iradierii au o viata foarte scurta, de
ordinul a 10-6 dintr-o secunda, totusi ei au posibilitatea
de a intra intr-o serie de reactii chimice cu substantele din materia
iradiata, determinand aparitia unor compusi noi.
Dintre
radiatiile ionizante, cele mai puternice sunt radiatiile
electromagnetice si in special razele gamma si razele Röntgen, care
au o lungime de unda mica si putere foarte mare de
penetratie.
Razele
gamma, care iau nastere in cursul dezintegrarii radioactive,
au lungimea de unda cea mai mica, cuprinsa intre 0.005 si
1.4 Å. Cea mai cunoscuta sursa de radiatii gamma este cobaltul
radioactiv (60Co).
Razele
Röntgen (X) sunt radiatii electronomagnetice cu actiune
similara razelor gamma, avand o lungime de unda cuprinsa intre
0.06 si 100 Å si o energie de activare de 0.01-0.1 MeV. Puterea lor de
penetratie este mai mica decat aceea a radiatiilor gamma, dar
suficient de mare pentru a provoca o ionizare puternica.
Tot
din grupa radiatiilor ionizante fac parte radiatiile corpusculare cum sunt razele a
(nuclei de heliu), razele b (electroni), protonii (nuclei de hidrogen), neutronii si diverse
particule grele, emise de asemenea in timpul dezintegrarii radioactive a
unor elemente.
Razele
alfa (a sunt formate din nuclei de
heliu, alcatuiti fiecare din doi protoni, cu o masa de circa
7.000 de ori mai mare decat electronul, ceea ce le confera o putere de
penetratie foarte mica. Distanta maxima de patrundere
a particulelor alfa emise de exemplu de radium este de aproximativ 4 cm in aer
si aproximativ 0.07 mm in tesuturile biologice, ceea ce face ca utilizarea
lor in lucrarile de mutageneza sa fie limitata.
Razele
beta (b reprezinta un flux de
electroni expulzati de nucleele atomice cu o viteza de 10.000-300.000
km/sec, avand o masa mica si o putere penetranta de circa
200 de ori mai mare ca a razelor alfa. Sursele cele mai cunoscute de raze beta
ce pot induce mutatii sunt izotopii radioactivi ai fosforului (32P)
sau sulfului (35S).
Neutronii
sunt particule elementare neutre emise in cursul unor reactii
nucleare, in special in reactiile de dezintegrare a nucleului de uraniu
si de plutoniu. Fiind lipsiti de sarcina electrica ei sunt
atrasi sau respinsi de
electroni sau nuclei materiei prin care trec.
In
functie de energia pe care o poseda, neutronii pot fi rapizi (cu
energii mai mari de 0.5 MeV), cu o viteza medie (avand o energie care
variaza intre 0.5 MeV si 1 KeV), lenti (cu energie cuprinsa intre 1 KeV si
1 eV) si termici (cu energie sub
0.1 eV).
In urma ciocnirilor cu nucleii
atomici ai materiei pe care o strabat, neutronii incep fie sa emita
protoni (ca in cazul iradierii cu neutroni rapizi), fie pot patrunde in
nucleu, fiind capturati de acesta (ca in cazul iradierii cu neutroni
lenti sau termici). In ambele situatii, iradierea cu neutroni duce la
producerea unui numar de particule incarcate electric si ioni,
care genereaza reactii chimice noi, caracteristice materialelor
ionizate.
Masurarea radiatiei. Unitatea de masura folosita curent pentru
radiatiile ionizante este rad-ul (radiation absorbed dose), care
reprezinta cantitatea de energie absorbita de 1 gram de materie vie
sau nevie. In paralel cu rad-ul (R) este frecvent folosita o
alta unitate de masura, respectiv gray-ul (Gy). Cele doua moduri de cuantificare a dozelor de
iradiere sunt compatibile, intrucat 1kR (1.000 razi) este echivalentul a 100 Gy.
Pentru studiul
efectelor radiatiilor asupra organismului uman se foloseste rem-ul
(röntgen equivalent man). Aceasta unitate de masura este o
varianta a röntgen-ului (r). Un röntgen este egal cu cantitatea de
radiatii capabile sa produca 2.08 x 109 perechi de
ioni intr-un cm3 de aer la 0°C si la presiunea atmosferica
de 760 mm Hg. Cantitatea de radiatii corespunzatoare unei
unitati röntgen este capabila sa provoace intr-un
tesut viu formarea unei cantitati de ioni de aproximativ 1.000
de ori mai mare, respectiv 2,08 x 1012.
Pentru
masurarea radiatiei neutronilor rapizi se foloseste ca unitate
de masura echivalentul fizic
röntgen, care reprezinta energia folosita pentru ionizare,
echivalenta cu energia absorbita de 1 g de apa iradiata cu
1 r de raze X. Masurarea dozelor de neutroni termici se face direct, prin
determinarea cantitatii de neutroni cu care este bombardata
intr-o secunda o suprafata de 1 cm2 din materialul
iradiat (Nt/cm2/s).
Radiosensibilitatea organismelor
Diferitele organisme
prezinta diferente considerabile in ce priveste
radiosensibilitatea. Pentru masurarea
radiosensibilitatii se folosesc notiunile de doza
letala 100 (DL 100) si doza letala 50 (DL 50), care
reprezinta doza la care, intr-un interval de 30 zile, mor toti
subiectii supusi iradierii, sau 50%.
Efectele
radiatiilor la nivel celular sunt foarte variate, ele determinand incetinirea sau blocarea diviziunilor
mitotice, pierderea definitiva a capacitatii de diviziune
neinsotita de moartea celulelor (sterilizarea celulelor), moartea
celulelor dupa mai multe ore de la iradiere (fara ca ele sa
mai intre in diviziune mitotica) sau, in cazul dozelor foarte mari de
radiatii (iradiere acuta), moartea instantanee a celulelor.
Indiferent de tipul
de iradiere, respectiv cronica (de intensitate scazuta si
durata lunga) sau acuta (de intensitate mare si durata
scurta), cele mai afectate componente ale celulei sunt nucleul si
cromozomii, deci materialul genetic.
Variatia
radiosensibilitatii la unele organisme vegetale si animale.
|
Tipul
de
organisme
|
Specia
|
DL 50
|
Plante
|
Lynum
ussitatisimum (in)
Lycopersicum
aesculentum (tomate)
Brassica
sp. (varza, nap, etc)
Secale
cereale (secara)
Phaseolus
vulgaris (fasole)
Abies
alba (brad)
|
|
Animale
|
Paramecium
aurelia (parameci)
Drosophila melanogaster (musculita de otet)
Salamandra
salamandra (salamandra)
Ratus
sp. (sobolan)
Rana
ridibunda (broasca)
Mus
musculus (soarece)
Canis
familiaris (caine)
|
|
|
Om
|
|
|
In general, frecventa mutatiilor creste proportional
cu doza de iradiere. De exemplu, la Antirrhinum majus frecventa
mutatiilor la 400 R este de 1.24%, iar la doza de 800 R creste
pana la 4.41%. In mod similar, la Drosophila
melanogaster frecventa mutatiilor letale la o doza
integrata de iradiere de 750-770 R este de 2.7%, iar la doza de 9.000 R (9
kR) atinge 18.3%.



Varietati
de Antirrhinum majus (stanga) si
mutante: plena (centru)
si
deficiens (dreapta)
Efectele genetice ale radiatiilor
Sub influenta radiatiilor
ionizante se pot produce ruperi ale cromozomilor, acestea fiind urmate cel mai
adesea de restructurari
cromozomiale.
Ca
urmare a unei singure rupturi cromozomiale, apare un fragment acentric, care de
obicei are tendinta de a se uni cu capatul liber al cromozomului care
a fost afectat de rupere. Daca prin aceasta
reunire, care poarta denumirea de restitutie,
secventa normala a genelor nu este alterata, cromozomul va fi
normal atat structural cat si functional.
Un
tip de modificare structurala a cromozomilor care se poate produce ca
urmare a radiatiilor ionizante este deletia,
caz in care ruperea nu este urmata de reunirea fragmentului acentric cu
restul cromozomului. Efectele genetice ale deletiei depind de marimea
fragmentului pierdut si de importanta genelor situate pe el.


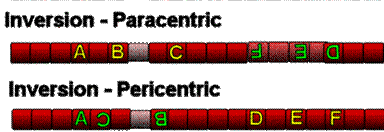
Daca ruperea unui
cromozom este urmata de reunire, dar inainte de reunire fragmentul
acentric face o rotatie de 180s, cromozomul va fi modificat din punct de
vedere structural, in sensul schimbarii ordinii locilor. Acest tip de
modificare poarta denumirea de inversie
terminala sau paracentrica.
In cazul in care cromozomul sufera doua ruperi, cu producerea a trei
segmente, dintre care numai segmentul care poarta centromerul isi
schimba orientarea, modificarea poarta denumirea de inversie pericentrica.

Alteori, un astfel de fragment se poate suda la capatul unui cromozom
neomolog determinand o translocatie
terminala.
In
cazul producerii de ruperi simultane in doi cromozomi neomologi poate avea loc
schimbul reciproc de fragmente acentrice intre ei, acest tip de restructurare
intracromozomiala fiind denumita translocatie
reciproca, care, in general, are o frecventa mai mare decat translocatia
terminala.
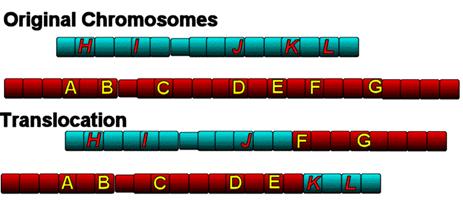
Daca
fragmentele translocate au centromeri, se vor forma cromozomi dicentrici sau
policentrici.
Prin
sudarea fragmentului acentric la cromozomul omolog se produce un alt tip de
restructurare intracromozomiala denumita duplicatie.
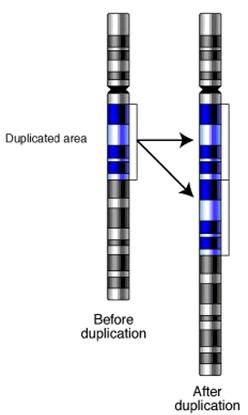
In functie de
modificarea sau nu a ordinii genelor inainte de sudura, aceasta poate fi duplicatie in tandem, cand sudura
nu schimba ordinea genelor, sau duplicatie
in tandem invers, cand ordinea initiala a genelor este
schimbata ca urmare a rotirii fragmentului acentric inainte de
sudura.
Duplicatia in
tandem invers poate avea efecte genetice majore, intrucat prin efectul de
pozitie poate fi afectata expresia caracterelor codificate de genele
afectate de restructurarea intracromozomiala.
Duplicatiile au o
frecventa mai mare decat deletiile, dar spre deosebire de
acestea sunt foarte rare cazurile in care au efect letal.
Fragmentele
acentrice se pot suda si ele, formand cromozomi inelari, dar acestia
se vor pierde la urmatoarea diviziune celulara, ca urmare a lipsei
centromerului.
Adeseori,
cromozomii dicentrici sau policentrici formeaza punti in
anafaza, acest tip de aberatii cromozomiale determinand aparitia
de gameti nefunctionali.
Este un fapt incontestabil ca plantele cu cromozomi mari sunt mai
susceptibile la aparitia de ruperi ale cromozomilor sau cromatidelor,
decat cele cu cromozomi mici, deci au o radiosensibilitate mai ridicata.
Se considera insa ca aproximativ 90-95% dintre rupturile primare
se reunesc in configuratia initiala (proces denumit
restitutie) intr-un interval cuprins intre cateva minute si cateva
ore. Celelalte rupturi, care nu refac configuratia initiala, pot
evolua pe urmatoarele doua cai:
(a) pot
ramane deschise;
(b)
se pot suda "nelegitim" cu capetele
rupte ale altor cromozomi,
avand
ca rezultat producerea de translocatii.
Aberatiile
de primul tip apar ca simple rupturi detectabile in mitoza, iar
fragmentele de cromozomi fara centromer se pot pierde, caz in care
poate avea loc si o pierdere de informatie genetica.
Aberatiile de al doilea tip
presupun existenta simultana a doua rupturi deschise, aflate in
imediata apropiere.
Pe baza relatiei intre
frecventa aberatiilor si doza de iradiere s-a ajuns la concluzia
ca in cazul radiatiilor X si gamma, o singura
particula are o probabilitate mica de a produce doua rupturi,
fapt care este insa perfect posibil in cazul radiatiilor dens
ionizante.
Din aceasta cauza, in cazul radiatiilor X sau gamma,
existenta a doua rupturi deschise presupune producerea a doua
rupturi independente si de aceea frecventa aberatiilor de acest
tip este egala cu produsul frecventelor celor doua rupturi
simple pe care le presupune.
Asadar, daca frecventa aberatiilor de primul tip este
dependenta de doza, frecventa celor de al doilea tip este
functie de patratul dozei.
Pentru a explica aparitia unor astfel de aberatii cromozomiale
ca urmare a iradierii, exista doua ipoteze:
Ipoteza
rupturii primare considera ca efectul primar al unui agent
exogen este o ruptura cromatidica sau cromozomica la nivelul
unui cromozom interfazic. Ipoteza presupune ca, de obicei, capetele de
ruptura se reunesc pentru a restaura configuratia originala
(restitutie), dar pot exista cazuri in care capetele de ruptura pot
ramane deschise sau se pot suda cu capetele altei rupturi. Rezultatul unei astfel de fuziuni a capetelor de la diferite rupturi ale
cromatidelor duce la realizarea a diferite schimburi cromatidice.
Ipoteza
schimburilor considera ca efectul primar al iradierii nu este
o ruptura, ci un alt tip de leziune la nivelul cromozomului, care poate
reveni la starea normala sau poate
da nastere unui schimb. Cand astfel de leziuni coincid in timp si spatiu cate
doua, ele se pot transforma (intr-un stadiu ulterior) in schimb cromatidic.
Refacerea cromozomilor
Se considera ca leziunile
produse de iradierea cu agenti mutageni pot suferi un proces de
restructurare spontana, dar aceasta poate fi provocata si de
factori fizici, sau de anumite substante cum sunt unii aminoacizi sau
proteine, derivati purinici si pirimidinici, nucleoside si
nucleotide, diferite enzime, etc.
Ipotezele
referitoare la mecanismele moleculare ce stau la baza repararii leziunilor
cromozomice sau cromatidice si la reunirea cromozomilor rupti se
bazeaza fie pe replicarea ADN ca proces esential al restaurarii,
fie pe implicarea unor proteine (enzime) in refacerea cromozomilor, fie pe
ambele procese. In acest sens s-a demonstrat de altfel ca o serie de
enzime de tipul endonucleazelor, exonucleazelor, polimerazelor, etc., au un rol
foarte important in procesul de refacere a cromozomilor afectati de
iradiere.
Efectele genetice ale radiatiilor
ionizante sunt dependente de doza, debitul dozei, tipul radiatiei,
viteza diviziunii celulare (durata ciclului mitotic), numarul si
lungimea cromozomilor, concentratia oxigenului intracelular si
extracelular, eficienta mecanismelor de reparare moleculara a ADN,
reversibilitatea leziunilor cromozomiale.
Factori mutageni chimici
Gama substantelor chimice
capabile sa induca mutatii (genice, cromozomiale si
genomice) este foarte larga. Deoarece multe dintre aceste substante
au asupra cromozomilor efecte similare radiatiilor, provocand de exemplu
ruperi, au primit denumirea de substante
radiomimetice.
Substantele chimice pot induce
aberatii variate de tip cromozomial, cromatidic si subcromatidic. In
general, insa, substantele chimice mutagene induc aberatii de
tip cromatidic si numai putine substante (de exemplu
8-etoxicofeina si streptonigrina) produc aberatii de tip cromozomial
sau subcromatidic.
Agentii
chimici care produc rupturi si restructurari cromozomiale sunt
clasificati astfel:
a) precursori ai ADN si analogi ai bazelor azotate;
b) agenti alkilanti;
a) unele antibiotice;
b) alti factori mutageni
chimici
Efectul agentilor mutageni
chimici este dependent de durata de contact, concentratia,
si nu in ultimul rand de o serie de factori
fizici externi (pH, concentratia O2, temperatura, etc).
Agentii
mutageni chimici afecteaza mai ales interfaza
si profaza timpurie. Substantele
chimice cu efect in perioadele G1 sau S ale interfazei vor inhiba
replicarea cromozomilor prin actiunea lor asupra sintezei de ADN, pe cand
cele cu efect in G2 vor afecta numai formarea cromatidelor libere,
intrucat in acest stadiu ADN si cromozomii sunt deja replicati.
Unii agenti
chimici, cum sunt adenina, 5-fluordeoxiuridina, azaserina, aminopterina, etc.,
blocheaza sinteza ADN si a precursorilor sai.
Alti agenti, cum
sunt mitomicina-C, actinomicina-D, hidrazida maleica, agentii
alkilanti, etc., modifica proprietatile chimice si
fizice ale ADN.
Se considera ca
blocarea sintezei unei anumite baze azotate determina probabil
aparitia unor erori in procesul de imperechere normala a
nucleotidelor purinice si a celor pirimidinice in macromolecula de ADN.
Cel
mai puternic agent mutagen chimic s-a dovedit a fi antibioticul azaserina, care blocheaza sinteza bazelor azotate purinice,
acesta fiind, in acelasi timp, si un
puternic alkilant.
O alta categorie de
substante chimice cu efect mutagen este aceea care include derivatii
halogenati ai benzenului si toluenului, aminopurinele, cafeina,
teofilina si teobromina, care inhiba citochineza la plante, avand ca
efect inhibarea diviziunii celulare fara a afecta diviziunea cromozomului
sau respectiv a nucleului.
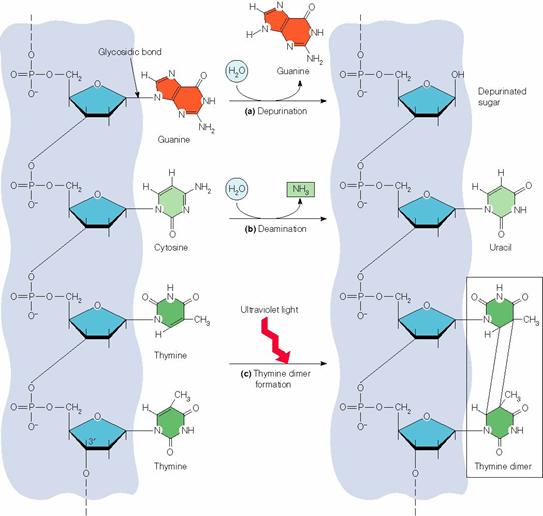
Cai de modificare a ADN
sub actiunea unor agenti chimici mutageni
O categorie distincta
de agenti chimici mutageni este aceea a
substantelor care actioneaza asupra fusului de diviziune.
Inhibarea formarii fusului are ca efect oprirea diviziunii celulei,
fara a fi insa afectati cromozomii, care isi
continua diviziunea in mod normal.
Precursori ai ADN si analogi ai bazelor azotate
Din aceasta
grupa fac parte un numar relativ mare de substante chimice cu
efect de rupere a cromozomilor si care induc mutatii, cum sunt:
adenina, dezoxiadenozina, 5-fluordezoxiuridina, 5-bromdezoxiuridina, 5-clordezoxiuridina, 5-iododezoxiuridina,
citozina arabinozida, oxipurinele N-metilate (teobromina, cafeina,
8-etoxicafeina, teofilina, acidul 1,3,7,9-tetrametiluric), 2-aminopurina,
2-6-diaminopurina, 5-bromuracilul, 5-cloruracilul, 5-ioduracilul,
5-floruracilul, etc.
Agenti alkilanti
Din grupul
agentilor alkilanti cu actiune mutagena fac parte numeroase
substante chimice, printre care iperita, dietilsulfatul, epoxizii, b-propiolactona,
azaserina, etc.
Modul de actiune al agentilor alkilanti. In general, desi exista mai multe ipoteze privind procesele
care duc la aparitia aberatiilor cromozomiale, se considera
ca acestea sunt cel mai probabil rezultatul sensibilitatii
foarte mari la alkilare a ADN. Agentii alkilanti actioneaza
numai in perioada S a ciclului mitotic, deoarece in celelalte perioade ADN
cromozomial este protejat de alte substante.
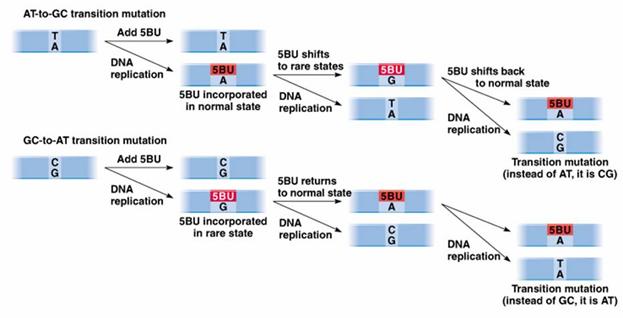
Actiunea
mutagena a 5-bromuracilului (5BU)
Sub actiunea
agentilor alkilanti, ADN este denaturat, devenind monocatenar.
Fenomenul de alkilare a ADN continua apoi prin separarea bazelor purinice
alkilate de lantul format de zaharuri si radicali fosforici. Ca urmare are loc ruperea lantului polinucleotidic
al ADN.
Antibiotice cu efect mutagen
Este un fapt bine
cunoscut ca mitomicina C, un antibiotic produs de ciuperca Streptomyces caespitotus, induce
aberatii cromozomiale cu o frecventa apreciabila, atat la
plante, cat si la animale. Acest antibiotic are un puternic efect
inhibitor asupra sintezei de ADN, fara a influenta marcant
sinteza ARN si a proteinelor.
Mitomicina C induce de
asemenea degradarea ADN si actioneaza ca un agent alkilant,
afectand preferential regiunea constrictiilor secundare.
Alti factori mutageni chimici
In aceasta categorie de factori mutageni este inclusa o
multitudine de substante chimice al caror mecanism de actiune
asupra acizilor nucleici este foarte variat si in mare masura
necunoscut. Printre aceste substante se numara acidul nitros,
hidroxilamina, hidrazina, clorura de mangan, peroxidul de hidrogen, unele
metale grele, unii alcaloizi, unii coloranti (de exemplu, acridin orange),
etc.
Acidul nitros (HNO2)
este unul dintre agentii mutageni al carui efect este cunoscut de
foarte mult timp, dar al carui mod de actiune la nivelul acizilor
nucleici a fost descoperit abia in urma cu 3 decenii.
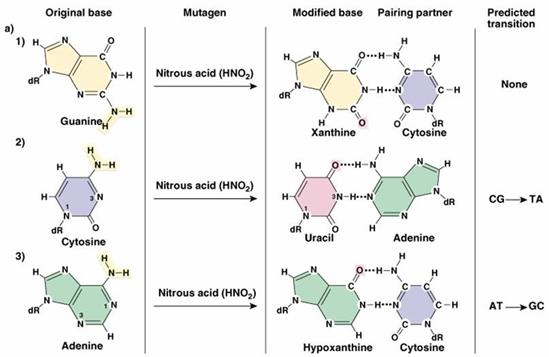
Producerea
de mutatii la nivelul macromoleculei de ADN
sub actiunea acidului nitros
Acidul nitros este
capabil sa induca modificari atat in macromolecula de ADN, cat
si in cea de ARN, mecanismul sau molecular de actiune constand
in dezaminarea unor baze azotate, ceea ce determina transformarea adeninei
in hipoxantina, a citozinei in uracil si a guaninei in xantina.
Mecanismul
molecular al mutatiilor
Cauzele care duc la aparitia
mutatiilor pot fi urmatoarele:
modificarea structurii
macromoleculei de ADN;
deletia sau aditia
uneia sau mai multor nucleotide in macromolecula de ADN;
substitutia uneia sau
mai multor nucleotide in macromolecula de ADN;
inversia unei secvente de
nucleotide din macromolecula de ADN.
Sub
influenta diversilor factori mutageni se produc erori in procesul de
replicatie a acizilor nucleici, care duc la aparitia de mutatii.
Inlocuirea unei baze purinice in macromolecula de ADN sau ARN cu o alta
baza purinica (A G), sau a unei baze pirimidinice cu o alta baza
pirimidinica (T C)
este denumita tranzitie,
in timp ce schimbarea unei baze purinice cu o baza pirimidinica sau
invers (A T; A C; G T; G C) este denumita transversie.
O cauza a unor astfel de erori o
constituie existenta unor baze azotate sub forma tautomerica
rara, aparute de exemplu prin schimbarea pozitiei unui atom
de hidrogen.
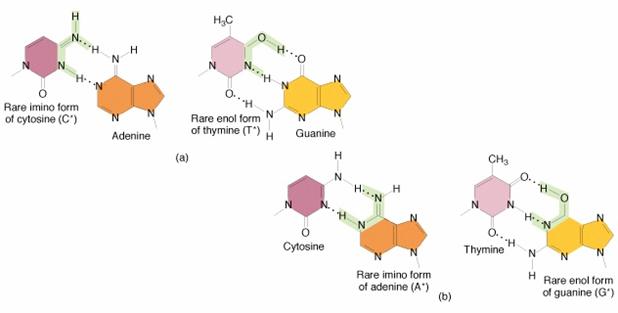
Forme
tautomerice rare ale bazelor azotate
In mod normal, adenina se
imperecheaza in lantul polinucleotidic cu timina, insa forma
tautomerica a adeninei se poate imperechea cu citozina, adica A-T
devine A-C. Tinand seama ca citozina are afinitate chimica pentru
guanina, la replicatia urmatoare a macromoleculei de ADN,
legatura A-C se transforma in G-C. Aceasta inseamna in fapt
ca perechea de baze azotate A-T este inlocuita de perechea de baze
G-C, fiind astfel modificata informatia genetica prin
mutatie.

Modificarea
unor nucleotide prin deaminare
Erorile de includere a
unor nucleotide in macromolecula de ADN si erorile de replicatie a
ADN constituie, la nivel molecular, mecanismele de baza care
realizeaza modificarea informatiei genetice si respectiv
aparitia procesului mutational.
Cercetarile
privind mecanismul molecular al procesului de mutageneza au aratat
asadar ca factorii mutageni actioneaza, de obicei, la nivelul acizilor nucleici, determinand
consecinte genetice importante. Modul de actiune al agentilor
mutageni este foarte variat, acestia provocand diferite modificari,
cum sunt inlocuirea unor nucleotide, aditia sau deletia de
nucleotide, rupturi sau chiar depolimerizari ale macromoleculei de ADN,
etc. O parte dintre aceste modificari se stabilizeaza in procesul
replicatiei acizilor nucleici, fapt ce determina modificari
structurale in macromoleculele de ADN si ARN. Ca urmare, apar
modificari ale informatiei genetice si respectiv ale procesului
de biosinteza a proteinelor.
Mutatiile
la nivelul macromoleculelor de ADN nu sunt intotdeauna reflectate in
schimbari ale secventei aminoacizilor, din cauza codului genetic
degenerat. De exemplu, codonii UAU si UAC codifica acelasi
aminoacid, si anume tirozina. Ca urmare, schimbarea nucleotidei ce
contine uracil (U) cu o nucleotida ce contine citozina (C)
in pozitia a treia a codonului nu are ca efect substituirea (inlocuirea)
aminoacidului respectiv.
Dintre
cei 64 de codoni, un numar de 3
(UAA, UAG, UGA) servesc pentru marcarea terminarii secventei de
aminoacizi a unei catene polipeptidice.
O mutatie a acestor 3 codoni nu
are efecte asupra secventei de aminoacizi si ca urmare o astfel de
mutatie a fost denumita mutatie
nonsens.
Daca insa are loc o
mutatie care determina transformarea intr-un codon terminal a unuia
dintre ceilalti 61 de codoni, aceasta poate avea ca efect incheierea prematura a secventei de
aminoacizi si deci formarea unei catene polipeptidice mai scurte.
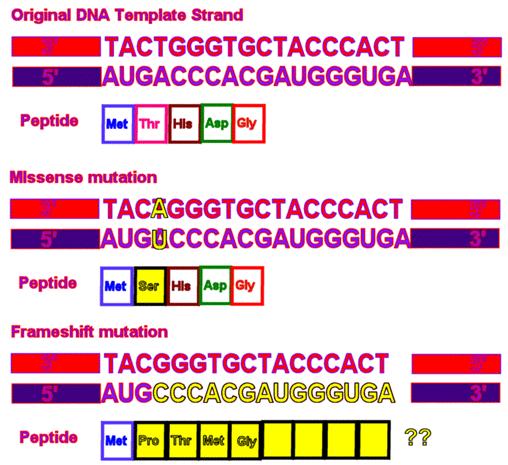
Efecte
ale mutatiilor produse la nivelul secventelor de nucleotide
Mutatia
non-sens
Mutatiile care afecteaza
unul dintre cei 61 de codoni si determina de obicei inlocuirea unui
aminoacid cu altul, se numesc mutatii
cu sens gresit. Datorita fenomenului colinearitatii,
mutatia unui codon dintr-o anumita pozitie din secventa de
nucleotide are ca rezultat inlocuirea unui aminoacid cu altul in pozitia
corespunzatoare din catena polipeptidica.
Mutatiile care afecteaza codonii, dar care nu duc la inlocuirea
unui aminoacid cu altul, se mai numesc mutatii sinonime sau mutatii neutre.
PRODUCEREA DE MUTATII SI RECOMBINARI
GENETICE
CA REZULTAT AL ACTIVITATII ELEMENTELOR
GENETICE TRANSPOZABILE
In genomul multor specii de
microorganisme, plante si animale, sunt prezente copii multiple ale unor
secvente de ADN care au capacitatea de a-si schimba pozitia
dintr-un locus in altul. Intrucat aceste secvente de ADN, care au primit
denumirea de elemente genetice
transpozabile sau transpozoni,
pot fi integrate in noul locus in cadrul genomului prin alte mecanisme decat
omologia secventelor de nucleotide, ele pot determina recombinarea genetica nelegitima. De asemenea,
elementele genetice transpozabile pot cauza mutatii, modificari in
reglajul genetic al unor gene, activarea sau inactivarea unor gene si a
celor adiacente locusului de integrare, restructurari cromozomiale (deletii,
inversii, duplicatii, translocatii).
Pana in prezent au
fost bine descrise 4 categorii de transpozoni:
1) elemente de control,
din care au fost deja descoperite 8 familii;
2) elemente de tip
retroviral;
3) Mu (abreviere pentru mutator);
4) insertii
biologic necaracterizate, cu structura similara
transpozonilor.
Elementele de control
pot fi, la randul lor, grupate in doua clase:
- prima clasa include
elementele autonome, care
contin toata informatia necesara pentru propria lor
transpozitie, dar a caror activitate (in ciuda autonomiei) poate fi
afectata atat de fondul genotipic cat si de factorii de mediu;
- a doua clasa
contine elementele neautonome,
care pot fi mobilizate doar in prezenta (oriunde in genom) unui element
activ inrudit. Astfel, fragmentul inactiv, mobilizat, si activatorul
formeaza un sistem dublu-element.
Cel mai bine cunoscut
sistem de acest fel este Ac-Ds. Majoritatea elementelor Ds sunt considerate a fi elemente Ac degenerate, in sensul ca sunt
inrudite cu elementele active Ac, dar
au suferit mutatii si/sau deletii care le-au inactivat
functia de transpozitie. Totusi, multe elemente Ds sunt omologe cu elementele active Ac doar in secventele terminale
repetate, care sunt tinte necesare in procesul de transpozitie.
Astfel, un membru neautonom al unei familii dublu-element poate fi construit
din segmente terminale adecvate care definesc capetele fragmentelor de
mobilizat, separate de orice secventa.
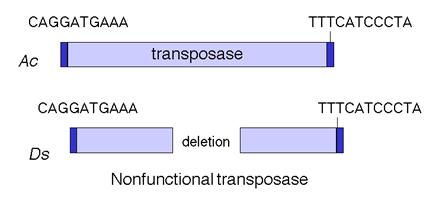
Elementele
genetice transpozabile Ac si Ds
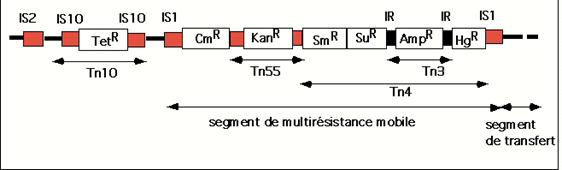
Elemente
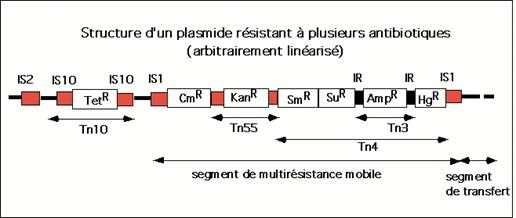
genetice transpozabile (transpozoni) complexe, continand gene pentru
rezistenta la antibiotice (Tn10; Tn55) si metale grele (Tn3)
Limita superioara
de marime a fragmentelor care pot fi mobilizate de transpozoni nu a fost
inca determinata, dar se apreciaza ca marimea acestora
poate fi de ordinul sutelor de kb.
Secventa de baze
pentru integrare pare sa fie in mod esential randomizata, dar
relatia dintre situsul de origine al elementului si situsul in care
se transpozeaza nu este in mod necesar randomizata. Astfel, Ac se deplaseaza mult mai frecvent
in pozitii apropiate celei de origine, decat in pozitii
indepartate si rareori se transpozeaza intre cromozomi. Ac este in mod obisnuit prezent in
genom intr-un numar redus de copii, asa incat este relativ simplu de
urmarit transpozitia, atat in termeni genetici cat si
moleculari. In fapt, frecventa de transpozitie este cu atat mai
redusa cu cat numarul de copii ale Ac este mai ridicat. In mod contrastant, Mu pare sa se deplaseze randomizat
in interiorul genomului, fara a fi limitat la transpozitii
scurte. Fagul Mu este capabil sa
se insereze in diverse locusuri in cromozomul bacterian, provocand o gama
larga de mutatii diferite. Ulterior s-a descoperit ca acesta
este de fapt un transpozon, care poate sa existe si sub forma unui
virus temperat. Capacitatea de replicare a fagului Mu este asociata cu cea de transpozitie, ceea ce poate
determina o serie remarcabila de rearanjamente cromozomiale, cum sunt
fuziunea intre doua molecule separate si autoreplicabile independent
(repliconi), deletia unor secvente de nucleotide, inversia unor
astfel de secvente, etc.
Se afirma ca
transpozitia poate avea sau nu efecte detectabile, insa insertia
unui element genetic transpozabil intr-o secventa de codificare
determina cu mare probabilitate inactivarea genei respective.
Totodata, s-a sugerat ca excizia imprecisa a elementelor
transpozabile poate genera rearanjamente (deletii, inversii) ale
secventelor cromozomale adiacente.

Mutatii
induse de activitatea transpozonilor la Petunia
Structura Moleculara a Transpozonilor
Cei
mai simpli transpozoni sunt cei descoperiti la bacterii
si cunoscuti acum sub denumirea de secvente de insertie sau elemente IS. Similar elementelor genetice transpozabile de la
eucariote, acesti transpozoni mici poseda secventele invers
repetate la capetele lor si codifica propria transpozaza (proteina enzima necesara pentru
producerea exciziei si realizarea transpozitiei); transpozonii
bacterieni codifica de asemenea una sau mai multe proteine implicate in
reglarea ratei transpozitiei.
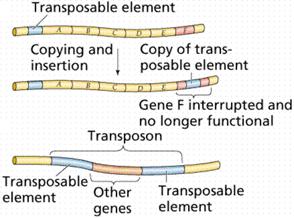
Transpozonii complecsi au dimensiuni mai mari, deoarece pe langa
genele necesare transpozitiei contin si una sau mai multe gene
mobile transpozabile, cum ar fi gene pentru rezistenta la antibiotice,
toxine, etc. Ca structura, un transpozon complex include un segment de ADN
mai mare, avand la extremitati cate un transpozon simplu (IS), iar in
partea centrala gena sau genele ce sunt transpozate.
Transpozonii sunt in mod
obisnuit desemnati prin abrevierea Tn
urmata de un numar (de exemplu Tn5). Cand trebuie sa
se faca referire la genele purtate de un astfel de element, se
foloseste si abrevierea pentru acestea. De exemplu Tn5(neo-r ble-r str-r) contine
genele pentru rezistenta la trei antibiotice diferite: neomicina
(kanamicina), bleomicina si streptomicina. Aceste gene reprezinta markeri, care fac usoara
detectarea transpozitiei unui element complex.
La
Escherichia coli sunt cunoscuti
mai multi transpozoni complecsi, cum sunt Tn3, Tn5, Tn10, etc. De
exemplu, transpozonul Tn3 este constituit dintr-un segment de ADN de 4.957 pb,
care are in partea centrala gena pentru rezistenta la
ampicilina, iar transpozonul Tn10 are o marime de 9.300 pb si
poseda gena pentru rezistenta la tetraciclina. Exista
si transpozoni complecsi mai mari, cum este transpozonul Tn2571, care
are o marime de 23.000 pb si poseda concomitent mai multe gene
de rezistenta (la streptomicina, cloramfenicol, sulfonamide,
etc).
Un tip particular de
elemente genetice transpozabile este acela al retrotranspozonilor, considerati
ca fiind inruditi (la origine) cu retrovirusurile care infecteaza
celulele eucariote. Retrovirusurile sunt virusuri care au dobandit capacitatea
de a se insera in genomul celular sub forma de provirusuri, iar sub
forma libera, adica de virioni sau particule virale, pot circula
de la o gazda la alta, spre deosebire de retrotranspozoni care pot circula numai in cadrul genomului
celular. Retrovirusurile si retrotranspozonii se aseamana in mod
deosebit prin faptul ca mobilitatea lor se realizeaza prin
intermediul ARN.
S-a demonstrat ca intre
retrovirusuri, provirusuri si elementele genetice transpozabile
exista similaritati structurale considerabile. Astfel, s-a observat ca, dupa
transpozitie, transpozonii sunt legati de o secventa
repetitiva inversata de pana la 1.500 pb. Secventa
repetitiva directa ia forma unei duplicatii de 4 pana la 12
pb la locusul de insertie in genom. Studiul locusurilor de insertie a
unor provirusuri si retrovirusuri a dus la constatarea ca
insertia in genomul celular se realizeaza in regiuni unde exista
o secventa repetitiva de 5 pb, alaturi de o
secventa inversata formata din 3 pb, secventele
respective fiind insa diferite pentru diferitele virusuri.
Se
considera ca in cursul procesului evolutiv, prin captarea de
catre transpozoni a unor gene pentru sinteza unor proteine (evident,
proteine pentru constructia capsidei), ele au dobandit capacitatea de a
circula nu numai in cadrul genomului, ci si extracelular, devenind
retrovirusuri. Mai
mult, s-a constatat ca transpozonii pot fi transportati de catre
virusuri peste barierele de specie, virusurile fiind de obicei capabile sa infecteze mai multe specii. Transpozonii pot deci
circula orizontal de la o specie la alta prin intermediul virusurilor, devenind un factor
important pentru procesul evolutiv, ca urmare a modificarilor importante
pe care le pot determina in materialul genetic al diverselor specii.
Influenta elementelor genetice transpozabile in modificarea
genomului se realizeaza prin sporirea variabilitatii genetice
si prin frecventa mai ridicata cu care organismele pot suferi
mutatii.
Tipuri de Transpozitie
De regula, transpozonii pot
migra si se pot insera in orice locus din genom, desi uneori
prefera sa se insereze in anumite secvente de nucleotide. S-a demonstrat existenta a trei moduri distincte de realizare a
transpozitiei elementelor genetice transpozabile:
1) transpozitie simpla;
2) transpozitie replicativa;
3) retrotranspozitie.
Prin
transpozitia simpla are
loc migrarea unui element genetic transpozabil intr-un alt locus din genom,
determinand astfel aparitia unui gol (gap)
in vechiul loc de insertie. Insertia sa in noul locus are ca rezultat
inducerea unor restructurari cromozomiale, inactivarea unei gene sau a
genelor adiacente, mutatii, etc.
Transpozitia replicativa se
realizeaza prin replicarea transpozonului, urmata
de migrarea copiei sale intr-un alt locus. Spre deosebire
de transpozitia simpla, transpozitia replicativa
determina o marire a numarului de copii ale transpozonului
respectiv per genom.
Retrotranspozitia se
realizeaza prin transcriptia transpozonului in ADN, urmata de
migratia in noul locus, unde cu ajutorul enzimei revers-transcriptaza se
produce transcriptia inversa in ADN. Sub aceasta forma,
retrotranspozonii sunt inserati in noul locus printr-un proces de
recombinare. Un exemplu de retrotranspozon este Ty de la Saccharomyces
cerevisiae, care se pare ca este inrudit cu retrovirusurile care au
genom ARN, cum este cazul virusului imunodeficientei dobandite umane
(HIV).
De
regula, mobilitatea transpozonilor este determinata de conditii
de stres (de exemplu iradierea cu radiatii ionizante sau neionizante,
socuri de temperatura, actiunea unor agenti chimici
mutageni), iar inserarea lor in genom se realizeaza probabilistic.
Intrucat
elementele genetice transpozabile pot fi integrate in noul locus in cadrul
genomului prin alte mecanisme decat omologia secventelor de nucleotide,
ele pot determina recombinarea
genetica nelegitima, pot cauza mutatii, modificari in
reglajul genetic al unor gene, activarea sau inactivarea unor gene si a
celor adiacente locusului de integrare, restructurari cromozomiale
(deletii, duplicatii, inversii, translocatii).
Elementele genetice transpozabile sunt asadar
implicate in restructurarea genomului, de activitatea lor fiind dependenta
intr-o masura importanta sporirea variabilitatii in
conditii de stres si adaptarea la conditii noi, nefavorabile, de
mediu.

